The rime (or rime ice, that can be divided in soft rime and hard rime, the former apparently very similar to hoarfrost) is a form of atmospheric precipitation that consists of a deposit of ice in the form of needles or flakes, which can occur in the presence of fog or high atmospheric humidity (above 90%) when the air temperature is below 0 °C. It appears as an opaque-white crystalline ice coating around solid surfaces, usually not very hard, brittle, and easily shaken off. The soft rime is formed by the passage of water from fog droplets in a supercooled state, i.e. remained in a liquid state even within temperatures beneath their solidification temperature, to ice. Supercooling occurs due to the small size of the water droplets – a very small drop has a surface tension such that it does not freeze – when the water is pure, i.e. without salts, and the cooling takes place in a state of quiet. Supercooling is an extremely unstable condition, and water in this state immediately solidifies as soon as it comes into contact with a solid object, a phenomenon known as gelicide. The ice crystal that is produced in this way has a different crystalline structure than classic ice crystals, and changes shape depending on the temperature of the supercooled water from which it derives. To form soft rime, the fog droplets must be very small, temperature must be low, there must be little or no ventilation and the latent heat of solidification must dissipate quickly. When these parameters change, other formations are produced, such as the hard rime, which is formed when the droplets of fog are thicker and the wind is stronger. Rime also differs from hoarfrost, since the latter does not derive from the supercooling of water droplets, but is formed by the frosting of steam on surfaces cooled due to heat loss by radiation during the night.
What could be of interest to lichen enthusiasts about this branch of meteorology? No idea. I was simply curious to find out whether rime and hoarfrost were synonyms, and Wikipedia gave me the answer. My interest in the rime, purely aesthetic, manifested itself yesterday, while I was browsing my photo archive to look for an appropriate photo for Christmas greetings, and I came across these images taken last November in Val Masino (Rhaetian Alps, Lombardy): Cladonias (obviously!) completely covered with sharp ice needles, a gift of the night frost of the Rhaetian valleys.
My sincerest wishes for a merry Christmas and a happy 2022 to all the blog followers!
The brand-new key to the Italian Cladoniaceae (Gheza & Nimis 2021) – fundamentally a crowd of Cladonia species, plus Pycnothelia papillaria and the mythological Pilophorus cereolus – has been online for a few hours and is now ready to be tested by lichenologists and amateurs.
This key is one of the numberless tesserae professor Nimis is assembling to create the monumental mosaic of the Italian Lichen Flora, the masterwork he started last year, which can be found in its own section of ITALIC and counts already 61 keys to single genera or families to date.
Since the previous attempt to regiment within a key the many species of Cladonia occurring in Italy had been performed by the writer (Gheza 2018), Prof Nimis kindly got me involved in this task. Actually, he carried out most of the effort: I just revised and readjusted few parts of a key that already worked very well in its original form.
About the key itself there is not much to say: it will speak for itself with anyone who will attempt to use it, hopefully successfully. Compared to mine, this key is more thrifty, and undoubtedly also more up-to-date: even if only few years have passed, several updates have been added at both floristic-biogeographical level (species previously unreported from Italy have been found, such as Cladonia corsicana, Cladonia subturgida and Cladonia zopfii) and taxonomic level (genetic studies suggested that some species should be ‘reabsorbed’ within others, e.g. Cladonia pseudopityrea seems to be just a skinny and haggard morphotype of the extremely variable Cladonia ramulosa, and similarly also the Italian specimens previously attributed to Cladonia prolifica should be brought back to that species). Furthermore, some species not yet reported from Italy have been included in the key, occurring not far from the borders of the country in habitats which are well represented also in Italy, since it is reasonable that they can also be found on Italian ground – and therefore they need to be included in the key, because otherwise it would be very difficult to realize whether you have found them, when identifying a papabile specimen.
There is no much more to say, just a warm invitation to everybody interested in Cladonia to test the key as soon as possible!
I can only conclude by expressing all my gratitude to Prof Nimis for allowing me to have a part – although an infinitesimal one if compared to the gargantuan amount of all the work he carried and will carry out – in this wonderful and monumental project of the Italian Lichen Flora. Such endeavor brightly demonstrates how Science made with great passion and also intended as a genuine sharing of knowledge (that is really vast, in his case) can really be a great gift for everyone. Vielen Dank Herr Professor, and full speed ahead!
Resilience…a word we heard often in the last year, at least here in Italy… Generally speaking, resilience is “the capacity of a system to cope with a change”. This term is used in different fields. In ecology, resilience is the capacity of an ecosystem or an organism to respond to a perturbation or disturbance by resisting damage and/or recovering, more or less quickly, after it.
Lichens can be quite resilient, and Cladonias are no less than the others. I have recently met a singular case of resilient Cladonia during an inspection in a very degradated grassland in the Ticino river valley. In that area, grasslands are encroached by the invasive tree Robinia pseudoacacia, and soil is frequently disturbed by the passage of quadrupeds…and bipeds as well. It can often happen to find cracked patches of Cladonias, especially of primary thalli of C. foliacea, C. polycarpoides or C. cariosa, as a result of heavy trampling. Sometimes, cushions of primary squamules are overturned, quite like old tortoises capsized on their shells, but, unlike tortoises, Cladonias are unable to regain their original position. What happens then? Obviously, the downturned part degradates and squamules die, because they are no longer reached by sunlight. But, if surrounding environmental conditions remain favorable, the former basal part, which is now placed on the top, can recover…and start producing new squamules! This is what I found in some old, overturned cushions of Cladonia polycarpoides: the downturned part was blackened and dead, but the upturned part was developing new, tiny squamules, and would soon regain its former appearance.
Cladonia…uh…finds a way.
A cracked patch of Cladonia polycarpoides…
…with the old squamules now dead or dying on the downturned upper side of the cushion…
…and new, tiny squamules developing on the upturned downside.
The so-called functional traits are the features of a living organism that play a role in driving its responses to the environmental factors interacting with it (Koch et al. 2019). Substantially, its biological, anatomical and morphological features. When using them for applied studies, functional traits can be described with quantitative values or just at a qualitative level. Traits of vascular plants are generally quantitative (stem lenght, seed viability, thickness of chlorophyll parenchyma, to name some of them), whereas traits of lichens are generally categorical. The second scenario allows to unite different species in the so-called functional groups, i.e. groups of species which have in common a certain trait. For example, in lichens functional groups are based on traits like thallus growth form (crustose, squamulose, foliose, fruticose, composite), photobiont type (chlorococcoid green alga, trentepohlioid green alga, cyanobacterium), reproduction type, etc. etc.
What is the difference between studying lichen communities’ traits and studying richness or composition, two parameters which have been more used until now? Functional traits can give us different information. Species are unique, whereas traits can group more than one species; and traits can differ between taxonomically or ecologically similar species. Therefore, studying traits can be important to understand why phylogenetically close species can behave in very different ways, or, vice versa, why phylogenetically distant species can show similar responses to the same environmental or disturbance factor(s). Between the 1990s and the 2010s, applied lichenological research used to investigate the effects of pollutants with the assessment of richness and composition of lichen communities, but the environmental emergencies currently acting on our planet – among which striking are the climate change and the loss/fragmentation/transformation of natural habitats – suggest that a more effective method to understand their impacts on lichen communities could be precisely to investigate their functional diversity, i.e. the diversity of their traits.
Investigating traits can be particularly revelatory when we deal with communities in which taxonomic diversity is low, but trait diversity is high. An exemplary case is represented by terricolous lichen communities in acidic dry grasslands of the western Po Plain: here, Cladonia is the only genus, sometimes with a high number of species, but anyway the belonging of all the species to a single genus somehow trivializes taxonomic diversity. But not the same is for functional diversity: in fact, Cladonia is a very diversified genus in terms of morphology (podetia can have very different shapes, and also primary squamules are variable), reproductive strategies and compounds produced by the secondary metabolism. And such variability had never been explored at the best of its potential, to date. This situation – lichen communities built-up by a single genus with a huge unexplored potential – took me to conceive a research that I developed together with some colleagues and that has been recently published on Microorganisms (Gheza et al. 2021).
Study area and study sites: acidic dry grasslands of the western Po Plain (Northern Italy) (from Gheza et al. 2021).
The focus of this research is on a very particular habitat: acidic dry grasslands, which are characterized by a precariousness due chiefly to vegetation dynamics, which leads to their disappearance. In fact, the typical composition of such grasslands includes pioneer plants, along with lichens and bryophytes, that can be found mainly in pioneer stages; when soil develops a layer of organic matter, vegetation evolves and the encroachment by more competitive plants causes the disappearance of pioneer ones…and of lichens and mosses! This would be a natural dynamics, that is however accelerated by the occurrence of some invasive plants which are very competitive and produce high amounts of organic matter, e.g. black locust (Robinia pseudoacacia). Vegetation dynamics is the main reason causing the need of an active managament of dry grasslands: management aims at maintaining pioneer conditions, in order to save the habitat requirements for pioneer plants and cryptogams. For this reason, the main environmental features considered in our research described the vegetation dynamics, to better elucidate how it influences functional traits of Cladonia. To do this, we used the vascular plants life forms [1], assuming that, in a grassland habitat, therophytes are the first colonizers and are dominant in pioneer stages, whereas hemicryptophytes and geophytes are dominant in intermediate-mature stages, and woody species – chamaephytes and phanerophytes – occur in mature stages and increase at increasing vegetation and soil maturity, sometimes also indicating ecotone situations. The other environmental features considered included local ones – soil pH, occurrence of disturbance by trampling and by an invasive Lagomorph that proved to be detrimental to lichens in this habitat (Gheza et al. 2018) – and climatic ones, i.e. mean annual temperature and annual rainfall. Such variables were put in relation, by means of statistical analyses, with three groups of functional traits purposely redefined with the explicit intention of highlighting the functional variability typical of genus Cladonia.
The main group of traits was the thallus growth form. Cladonia has a unique feature in the world of lichens: it has a composite thallus, built-up by the union of the so-called “primary thallus” and “secondary thallus”. The primary thallus is the basal part, made of little scales called squamules. The secondary thallus is the most morphologically variable part, made by structures called podetia, whose aim is to produce the apothecia. Podetia can have very different sizes and shapes across the species. In spite of this variability, previous literature assigned to Cladonia just two (foliose, fruticose) or three (foliose, fruticose with simple podetia, fruticose with branched podetia) growth forms. Considering the possible combinations between the two parts of the thallus and the morphological types of podetia, we introduced the use of six growth forms.
The six growth forms considered in this study, represented in scale; it can be easily noticed that also the average sizes of each form, aside of morphology, are very different.
The second group of traits was the reproduction strategy. Previous literature considered only if reproduction is mainly sexual (by means of apothecia in Cladonia) or asexual by some vegetative propagule (soredia are the most common in Cladonia). But the peculiar morphology of Cladonia allowed a further possibility, in addition: due to the composite thallus, in Cladonia pycnidia can occur either on primary squamules or on podetia, depending on the species. Therefore, the other element we took in account to describe the reproductive strategy was the position of pycnidia on the thallus. It could be assumed that species which produce pycnidia on primary squamules can do it faster than species which produce them on podetia, since squamules develop first; and therefore species with pycnidia on squamules could be faster and more effective colonizers of pioneer substrates/habitats.
The two descriptors of reproductive strategy: apothecia vs soredia (above) and pycnidia on squamules vs pycnidia on podetia (below).
The third group of traits included the secondary metabolites. Lichen compounds, which are produced exclusively by lichens, are widely studied due to their interesting pharmaceutical potential [2], but still largely unexplored under the standpoint of their ecological roles. And obviously the latter is far more intriguing to explore, for the ecologists. Among the most common metabolites in Cladonia we find atranorin, zeorin, and fumarprotocetraric, homosekikaic, rangiformic and usnic acids.
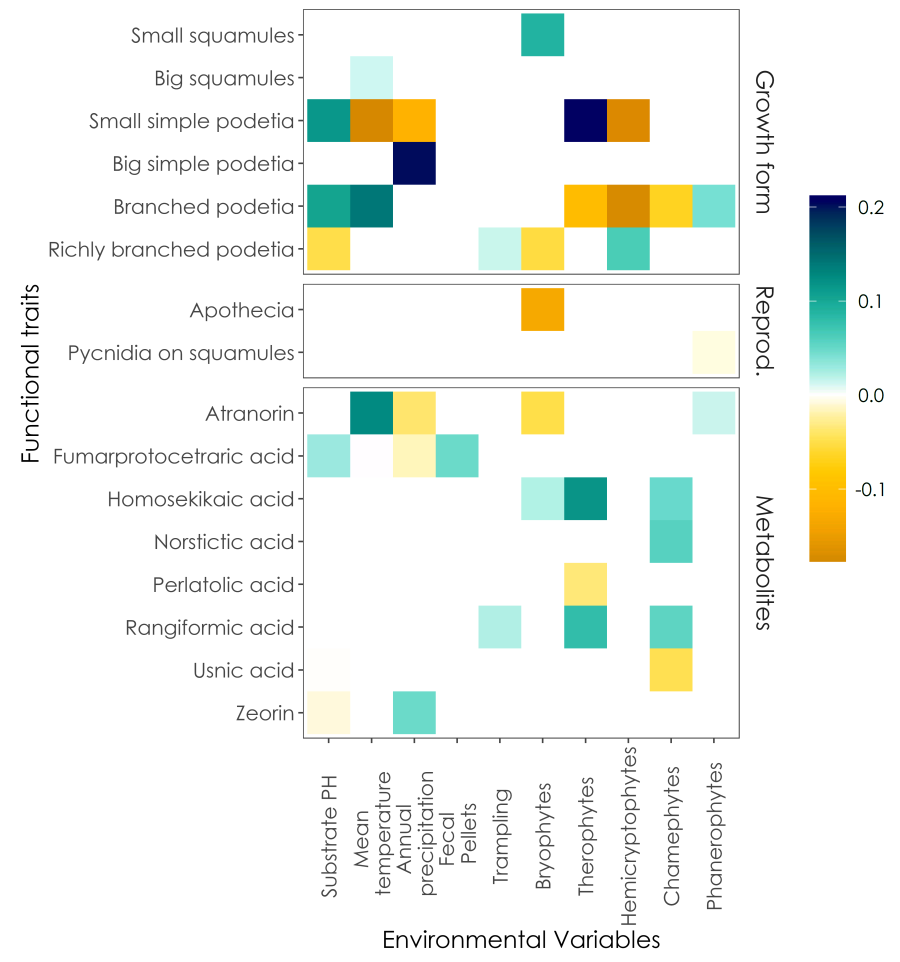
Surprisingly, the statistical analysis revealed more correlations than expected between the traits of Cladonia and the selected environmental variables. Furthermore, some of these correlations were very interesting and allowed to formulate some hypotheses that can help improving our understanding of trait-mediated interactions between lichens and the surrounding environment.
Significant correlations (coloured squares) between Cladonia functional traits and the considered environmental variables; positive correlations in cold colours, negative correlations in warm colours (from Gheza et al. 2021).
Many correlations were related to descriptors of vegetation dynamics, which revealed that species with small simple podetia and pycnidia on the primary squamules are positively related to pioneer stages and negatively to intermediate-mature stages, whereas an opposite pattern was evident for species with branched podetia, that were related to mature stages.
Interesting were also correlations disclosed between some metabolites and some environmental features, even more if considering that previous literature that investigated similar correlations is very scarce (one of the best is the detailed work by Zraik et al. 2018). Striking was the case of two metabolites with opposite correlations to what we can call a “canopy”, i.e. the cover of the vascular plants which produce a thicker cover (chamaephytes and phanerophytes, that can really be compared to what is for us a forest canopy, if seen from the perspective of organisms tall only few centimetres like lichens): usnic acid correlated negatively, whereas atranorin correlated positively. Previous literature suggested a photoprotection function for usnic acid (Farkas et al. 2020), which is therefore appropriate to find in species growing in exposed sites, and a role in facilitating the exploitation of low light intensities for atranorin (Rao & LeBlanc 1965), which is therefore appropriate to find in species that can grow under a moderate shading. In this case, our findings support interesting ecological roles already hypothesized by previous research.
I got also a valuable hint to deepen a question that has been in my mind for long: how is it possible that three species which differ so much in size, morphology and reproductive strategy, i.e. Cladonia cariosa, Cladonia rei and Cladonia rangiformis, have in common the fact of being widespread and often abundant also in disturbed and prohibitive habitats, and generally dominant in the lichen communities they occur within? The answer may lie within their metabolites: in fact, these three species contain homosekikaic and/or rangiformic acids, both of which correlated positively with all the dynamic stages of vegetation – pioneer and intermediate-mature. This suggests that the trait that drives such a wide distribution and ecological amplitude could well be the occurrence of one (or both) of these two metabolites. A better understanding of the ecological role of these two substances could perhaps help us to better understand the success of the species that produce them: a promising starting point for future research.
C. cariosa (left), C. rei (centre) and C. rangiformis (right) are the most successful species in this habitat, which often dominate the lichen vegetation in which they occur.
Finally, also climatic features correlated to some traits, and some of these correlations can be an alarm bell towards climate change. Most of these correlations gave a negative signal, but the most striking one is surprisingly positive. In fact, atranorin correlated positively with temperature and negatively with precipitation, suggesting that the modifications induced by global warming could foster species that produce such metabolite.
I am very satisfied for this study, not only because it allowed to deepen the diversity of Cladonia traits in a way that they really deserved, but also because it gave several interesting hints. The next step shall be to follow and deepen some of these hints…
References
Farkas E., Biró B., Szabó K., Veres K., Csintalan Z., Engel R. 2020. The amount of lichen secondary metabolites in Cladonia foliacea (Cladoniaceae, lichenised Ascomycota). Acta Botanica Hungarica 62: 33-48.
Gheza G., Assini S., Marini L., Nascimbene J. 2018. Impact of an invasive herbivore and human trampling on lichen-rich dry grasslands: soil-dependent response of multiple taxa. Science of the Total Environment 639: 633-639.
Koch N.M., Matos P., Branquinho C., Pinho P., Lucheta F., de Azevedo Martins S.M., Ferrao Vargas V.M. 2019. Selecting lichen functional traits as ecological indicators of the effects of urban environment. Science of the Total Environment 654: 705-713.
Rao D.N., LeBlanc F. 1965. A possible role of atranorin in the lichen thallus. The Bryologist 68: 284-289.
Zraik M., Booth T., Piercey-Normore M.D. 2018. Relationship between lichen species composition, secondary metabolites and soil pH, organic matter, and grain characteristics in Manitoba. Botany 96: 267-279.
Notes
[1] Plant life forms were introduced by the Danish botanist Christen Raunkiaer, and are therefore known also as “Raunkiaer System”. Life forms classify vascular plants by the mode in which they overcome the adverse season, considering as the main feature the position of the wintering buds. Hereinafter the definitions of the life forms mentioned in our research. Therophytes: annual herbs that survive the adverse season in the form of seed. Hemicryptophytes: biennial or perennial herbs and forbs with wintering buds at or near the soil surface. Geophytes: perennial herbs and forbs with subterranean resting buds within rhizomes or bulbs. Chamaephytes: perennial plants with a woody base with perennating buds close to the soil surface (between 5 and 25 cm from soil). Phanerophytes: perennial woody plants with perennating buds above 25 cm from the soil surface.
[2] Among the ecological roles attested for some of these metabolites, striking is the biocidal action, that can be directed towards different fronts: to control the proliferation of photobiont cells within the thallus, to fight competitors in the surrounding environment by means of allelopathy, to discourage herbivores to feed on thalli that contain them. Due to this activities, some of these metabolites are widely studied to exploit their antimicrobial, antifungal and even anticancer potential.
“Natura maxime miranda in minimis” (“Nature is greatest admired in the tiniest things”) is an expression attributed to Linnaeus – who in turn took it from Plinius the Old – that describes very well the wonder one feels when observing the marvellous and tiny complexity of organisms such as lichens.
Not all the lichens are microscopic, but it happens that also in groups characterized by conspicuous and showy species, like genus Cladonia, there can be also more discreet species, which risk to remain overlooked just because of their inconspicuous size.
Among these species we can find Cladonia peziziformis, a very tiny and often unnoticed species which is currently rare and threatened in the whole of Europe, to the point that it is protected in some Countries, e.g. in Great Britain, where it is considered a priority species (Chambers 2003) assessed as critically endangered (Woods & Coppins 2012). This lichen has lilliputian podetia, generally not taller than a half centimetre, and, since it grows directly on soil, it can be quite difficult to notice. One could therefore think that its putative rarity in Europe is due to this difficulty of detection, but instead it is actually rare and declining, mostly because it is closely bound to specific habitats which are declining as well, i.e. heathlands (Tønsberg & Øvstedal 1995). Until few years ago, in Italy this species had been reported only from Liguria, but recently it has been recorded in several sites of the western Po Plain (Piedmont and Lombardy) in which Calluna vulgaris-dominated heathlands and/or acidic dry grasslands still occur (Gheza 2018, 2020; Gheza et al. 2019a), which confirmed its bond to such endangered habitats.
Cladonia peziziformis
A similar species which can share the same habitat is Cladonia cariosa, that is however far more widespread in the whole of Italy and Europe. We could say that where we find C. peziziformis also C. cariosa is very likely to occur, but not vice-versa. So far, the most interesting sites where observe both the species together were located along the upper Sesia rivercourse (Gheza et al. 2019a; Gheza 2020), but unfortunately the anomalous flood of October 2020 buried under about a metre of pebbles and debris most of the dry grasslands located near the riverbed, causing a considerable decrease in the number of occurrence sites of both the species in the Sesia river valley. Which, of course, upset me.
Cladonia cariosa
Almost like a sort of ‘compensation’ from the karma, however, during one of my last inspections in the Ticino river valley on account of the Life Drylands project for which I am currently working, I accidentally found a new occurrence site of both the species in a small patch of acidic dry grassland.
The curious thing is that the last time I explored that grassland – right seven years ago, during the fieldwork for my MSc thesis – it was almost devoid of lichens! Just few thalli of the commonest species occurred – C. foliacea, C. rangiformis, C. rei. Instead, now 8 species of Cladonia occur, among which C. peziziformis and C. cariosa, which were absent seven years ago.
The grassland in which I have recently found Cladonia peziziformis within the Parco Ticino Lombardo (Lombardy Ticino Natural Park); even if it seems not exactly a biodiversity-rich habitat in its winter look studded with withered grass tufts, it is actually an important habitat for many organisms, not just for lichens.
Both these species had already been reported from the Ticino river valley (Gheza 2018; Gheza et al. 2019b); in spite of this, the discovery of this new site, considering also the relevant extent of the colonies, is surely an important information to know and take in account for future conservation planning.
Cladonia cariosa
As outlined above, Cladonia peziziformis and Cladonia cariosa are very similar and easy to mistake with each other, if one has only a scarce experience about. They have both capitate podetia which bear one or more wide brown apothecia, with a range of morphological variation which is enough to create some confusion in a newcomer. The most reliable feature to discriminate between the two species is the primary thallus: C. peziziformis has rounded or ear-shaped squamules with entire margin and procumbent posture, whereas C. cariosa has incised and generally erected squamules. Colours are slightly different as well – since also secondary metabolites differ between the two species – since C. peziziformis is more greenish, while C. cariosa is somehow more grayish. But this feature can be misleading, so it is always better to check carefully the primary thallus.
Which one is which one?!? (both the species are featured in this single shot) – In the dry grasslands on acidic sediments in the upper western Po Plain, these two species often occur together, and they characterize a particular lichen community which is typical of pioneer substrates (Gheza et al. 2019a).
A (not-so-)tiny satisfaction was also seeing that the post on the official Facebook profile of the Life Drylands project about this finding has been a great success, totalizing the highest number of ‘likes’ of all the contents ever posted on the page, and furthermore it has been shared by a considerable number of followers. I would have never expected such success for a post about tiny lichens! I really hope that also this can contribute to increase the awareness of the general public about the urgent necessity to protect and restore natural habitats as the only efficient way to protect also many rare, sensitive and fascinating organisms like these tiny lichens.
A rich terricolous lichen community featuring C. peziziformis, C. cariosa, C. polycarpoides and C. rei scattered amongst withered tufts of Armeria arenaria.
References
Chambers S.P. 2003. Site Dossiers for the UK BAP lichens Biatoridium monasteriense, Cladonia peziziformis, Graphina pauciloculata and Schismatomma graphidioides. CCW Contract Science Report N. 586.
Gheza G. 2018. Addenda to the lichen flora of the Ticino river valley (western Po Plain). Natural History Sciences 5 (2): 33-40.
Gheza G. 2020. I licheni terricoli degli ambienti aperti aridi della pianura piemontese. Rivista Piemontese di Storia Naturale 41: 147-155.
Gheza G., Barcella M., Assini S. 2019a. Terricolous lichen communities in Thero-Airion grasslands of the Po Plain (Northern Italy): syntaxonomy, ecology and conservation value. Tuexenia 39: 377-400.
Gheza G., Nicola S., Parco V., Assini S. 2019b. La diversità lichenica nella Valle del Ticino: conoscenze in continua evoluzione. 32° Convegno della Società Lichenologica Italiana, Bologna, 18-20 September 2019, poster presentation – Notiziario della Società Lichenologica Italiana 32: 56.
Tønsberg T., Øvstedal D.O. 1995. Cladonia peziziformis new to Norway from a burnt Calluna heath. Graphis scripta 7: 11-12.
Woods R.G., Coppins B.J. 2012. A Conservation Evaluation of British Lichens and Lichenicolous Fungi. Species Status 13. Joint Nature Conservation Committee, Peterborough.
On the top of the Old Stump there lived a family of Cladonias…
Quarantine due to the COVID-19 pandemic has reached most of the Northern Hemisphere in the last week. Here in Italy the lockdown lasts since the beginning of March, and even if I can do most of my work from home, I obviously miss very much going in the field (for fieldwork, but also for relaxation), especially because this would be the best period to do it.
Therefore, in my spare time I started to draw again – not only serious drawings, but also those frivolous cartoons I was well-known for during college years.
Some frivolity involves, of course, also my Cladonias, and right yesterday evening I had a flash in my mind about ‘living’ Cladonia characters, which I immediately had to transform into a quick sketch.
Do you know Cladonia sulphurina?
Its podetia have a worn appearance, being often crooked and deflated; cups have tattered and perforated edges and apothecia are usually flattenend, folded-up and deformed, not plain or globular as in most red-fruited species. Also the overall colour seems somewhat unhealthy, a ghostlike, almost phosphorescent yellowish-greenish which becomes a corpse-yellow in the herbarium.
It seems perfect to become the character of a cartoon or a comic strip! And I thought it was nice to portray them as a little family – a quarrelsome one, it would seem – inhabiting the ancestral family mansion on the top of an old stump, which is a typical substrate for the species.
This is just a concept for now, but, you known, quarantine is still long…
This family of Cladonia sulphurina has lived on the same old rotting stump for almost 50 years. Not once they complained for their sedentary lives, not once they tried to come down to go for a run in the understory. For 50 years. Yes, you’re right, now they look rather badly-off, but that’s the normal appearance of Cladonia sulphurina! #stayathome
Postscript – obviously I am very concerned for the situation, especially because I live in Lombardy, the most severely stricken area of Italy. I kept a light tone in this post, and avoided talking too much of it, but the situation is still dramatic. So, let’s really do it: stay safe, stay at home!
After running into this inspiration source, could I possibly have avoided launching a similar one with Cladonias!? Of course NOT!
So, I decided to illustrate the squamulose Cladonias occurring in the lowland dry habitats of the Po Plain.
This decision is due to more than one reason: they are species I know quite well now; they are often difficult to discern on the field (generally, to be 100% sure, TLC is needed), therefore a little scheme about them could be helping; and they occur in the Natura 2000 Habitats concerned by our Life Drylands project (btw: here’s the official website, launched few days ago).
For this last reason, at the bottom the chart you also find a little scheme of the most typical squamulose-Cladonia community of each Habitat.
If this seems a mess to you…well, I assure you that on the field it is far more worse!
There is no scale bar, but all the specimens are drawn proportionately between them, according to a ‘standard’ size based on my experience in the Po Plain.
Smaller figures indicate the habitus of each species in their natural habitat: some species form compact tufts or cushions (C. polycarpoides, C. strepsilis, C. caespiticia), some species usually form extended but flatter carpets (C. pulvinata), some species can do both (C. foliacea).
The pictures show a fertile specimen of Cladonia strepsilis, with very small (2-3 mm tall) podetia topped by small and ‘lumpy’ light brown apothecia.
The specimen comes from a cryptogam-rich heathland fragment developed on a shallow soil layer above a schist outcrop at about 900 m a.s.l. in the Orobic Alps. Here, C. strepsilis was associated with C. coccifera, C. chlorophaea, C. cervicornis, Pycnothelia papillaria and some mosses, in what is a typical heathland cryptogam community in northern Italy.
Fertile specimens of Cladonia strepsilis are uncommon. In this species podetia are rare, and when present they are often inconspicuous. They are often described as irregularly-shaped, and in fact they can have a wide morphological variability. The range of different and ‘irregular’ morphologies is due to the variable developmental pattern of the fungal meristem which is responsible of podetial development: “early ontogeny of the meristem includes combinations of splitting, enlarging, and budding, which give rise to mature podetia that are difficult to characterize” (Hammer 1999).
Podetia are usually small (<1 cm tall), turgid, fissurate, sometimes squamulose, with small branches bearing brown apothecia on the top, and “some podetia of C. strepsilis resemble primary squamules as a result of deflexed growth and elongation of the meristem tissue” (Hammer 1999).
Pictures of better developed C. strepsilis podetia can be found here, here and here.
Even if the ontogeny of the podetia of this species has been very well elucidated (Hammer 1999), the environmental factors which foster or hinder the development of podetia in this and other generally sterile Cladonias are still somewhat unclear. It would be interesting to better study them, which would be useful to better understand their ecology, and maybe to better plan conservation actions.
About C. strepsilis I have observed that thalli are usually sterile at the extremes of the altitudinal range (which are, in Northern Italy, the acidic dry grasslands in the western Po Plain and the cryptogam-rich grasslands in the alpine belt of siliceous Alps), and the few fertile ones I found were in sites located at average altitudes in the montane belt; but these observations are too circumscribed to infer reliable information. A wider study about the factors which drive the production of podetia in usually sterile Cladonias would be very interesting.
In conclusion of the XXXI meeting of the Italian Lichen Society in Pistoia (Tuscany), in September 2018, I took part to the final excursion which took place at Monte Ferrato, a huge ophiolite outcrop located in the outskirts of the city of Prato.
Ophiolite is a very interesting substrate for lichens (see the review by Favero-Longo et al. 2004), and I was curious to check which Cladonias would occur there, after having developed in the past years some knowledge on the Cladonias of the ophiolite outcrops of the province of Pavia (Lombardy).
The other peculiarity of the site chosen for the excursion was that the area had been investigated 80-90 years ago by the spouses Ettore Sambo (1884-1971) and Maria Cengia Sambo (1888-1939), who lived in Prato. Maria Cengia Sambo in particular is remembered as one of the few Italian lichenologists between the “Golden Age of Italian lichenology” (from the 1840s to the 1910s) and the “Reinassance of Italian lichenology” (from the late 1980s). However, her skills in identifying lichens are still controversial, and some of her identifications are considered unreliable – especially in some cases in which she proposed new taxa.
Both the Sambos investigated lichens on Monte Ferrato: Sambo (1927) under a floristic perspective, Cengia Sambo (1937) considering also the ecology of the lichens on the ultramafic substrate and the morpho-physiological modifications caused by it (e.g. “stenophyllous forms”, thalli with underdeveloped lobes due to the harsh conditions found on ophiolite substrates).
Overall, in their 2 papers (Sambo 1927; Cengia Sambo 1937) the Sambos reported 21 Cladonia taxa (Appendix 1), whereas during the SLI excursion on M. Ferrato I managed to find only 4 terricolous species (Appendix 3), which are more or less the same ones I collected on the ophiolites of the Lombardy Apennine – perhaps there is also a fifth species, but its primary thallus is so underdeveloped (or just young) that I wasn’t able to identify it.
Even if we investigated only a small area, and therefore we could have missed some species (since the Sambos explored a very larger extent of Monte Ferrato and neighbouring hills), some considerations can be done, especially in the light of the misidentifications of Cladonia specimens carried out by the Sambos (Appendix 2).
The first thing emerging from the two papers by the Sambos, about Cladonia, is a nomenclatural and taxonomic mess. For example, C. muricata, C. pungens and C. rangiformis are listed as different species, while nowadays they are all recognized just as synonyms of C. rangiformis; the same is for C. alcicornis and C. endiviaefolia, which both indicate C. convoluta – which has recently been synonymized with C. foliacea (Appendix 1).
The other problem is of biogeographical nature: the records of strictly (sub)alpine species such as C. amaurocraea, C. phyllophora, C. stellaris, C. uncialis, were already considered very dubious by Nimis (1993), and seemed very unlikely also to me. In spite of what stated by Cengia-Sambo about the “alpine species came down from the Alps unto Monte Ferrato”, environmental and climatic conditions were probably unadequate to those species also a century ago, and according to Occam’s Razor an erroneous identification is the most reliable explanation. It could easily be, e.g., that the reindeer lichens reported by the Sambos (C. rangiferina, C. stellaris) were instead C. portentosa, which still occurs in the hills of Tuscany.
We were able to ascertain [some of] the erroneous identifications, because during the writing of the paper (Munzi et al. 2019), Luca Di Nuzzo went to the ECI (Central Italian Herbarium) of Florence, where some of the lichen collections of the Sambos are still preserved, in order to check the real identity of the available specimens (Appendix 2).
It was surprising that the Sambos misidentified also easy and common species. We must consider that at that time things were more difficult for lichenologists, especially for the few Italian ones, who were isolated from the Central-European lichenology; however, mistakes like attributing to C. amaurocraea and C. uncialis two specimens of C. rangiformis and a specimen of C. furcata to C. convoluta are quite surprising.
Among those specimens, that of C. portentosa was recorded under “C. rangiformis” and there are no specimens reported under “C. rangiferina” nor “C. stellaris“, so the question about the real identity of the reindeer lichens found by the Sambos remains.
Finally, it is strange that the Sambos did not report C. symphycarpa, which is quite common on basic substrates and also on ophiolites in the Northern Apennine (pers. obs.); but maybe they found it and just reported it as a “strongly stenophyllous Cladonia foliacea“…
This research was useful to better understand the real identity of the Cladonias reported from Monte Ferrato. Also for other lichen genera interesting observations were made (see Munzi et al. 2019).
This points out the importance of checking historical records which appear dubious, that is possible only when matching specimens are stored in historical herbaria. So, once again, the importance of herbaria is highlighted by this story.
The jolly group of lichenologists at the excursion on Monte Ferrato at the end of the XXXI meeting of the Italian Lichen Society in Pistoia (Tuscany).
Appendix 2 Corrections to the erroneous Cladonia identifications by the Sambos (thanks to Luca Di Nuzzo):
“Cladonia amaurocraea” > Cladonia rangiformis
“Cladonia degenerans” [= C. phyllophora] > Cladonia rangiformis
“Cladonia endiviaefolia” [= C. foliacea f. convoluta] > Cladonia furcata
“Cladonia rangiformis” > Cladonia portentosa
“Cladonia squamosa” > Cladonia cervicornis
“Cladonia uncialis” > Cladonia rangiformis
After all of this mess, I wonder about the cited species of which no specimens were available in ECI…
Appendix 3 Cladonia taxa observed during the excursion of the XXXI meeting of the Italian Lichen Society (Munzi et al. 2019): Cladonia coniocraea Cladonia foliacea f. convoluta Cladonia pyxidata Cladonia rangiformis Cladonia symphycarpa
References Cengia-Sambo M. 1937. Osservazioni lichenologiche sul gruppo del M. Ferrato. Nuovo Giornale Botanico Italiano n.s. 44: 295-311.
Favero-Longo S.E., Isocrono D., Piervittori R. 2004. Lichens and ultramafic rocks: a review. The Lichenologist 36 (6): 391-404.
Munzi S., Benesperi R., Bianchi E., Brackel W.v., Di Nuzzo L., Favero-Longo S.E., Gheza G., Giordani P., Matteucci E., Paoli L., Tonon C. 2019. Sulle orme dei Sambo – escursione del XXXI congresso della Società Lichenologica Italiana al Monteferrato. Notiziario della Società Lichenologica Italiana 32: 99-108.
Nimis P.L. 1993. The lichens of Italy: an annotated catalogue. Monografie XII. Torino, Museo Regionale di Scienze Naturali di Torino. 897 pp. Sambo E. 1927. I licheni del M. Ferrato (Toscana). Nuovo Giornale Botanico Italiano n.s. 34: 333-358.
A great satisfaction, not only for the prestige, but mostly because this LIFE shall give us the opportunity to contribute concretely to the improvement of very important Habitats – which are particularly worthy to me, due to their lichen richness – dry grasslands and Calluna heathlands.
The poetic landscape of a “Heide” near the Malpensa Airport
The lichen-rich dry Habitats targeted by LIFE DRYLANDS are very important, in particular in the Po Plain, heavily impacted by human activity, where they are almost disappeared nowadays. Their importance lies mainly – but not only! – in the biodiversity they host, since they are home to very particular species which can live only in these dry and semidry habitats and cannot live anywhere else. For example, some rare terricolous lichens are found in Northern Italy only in these habitats. But they are not only plant and cryptogam species, but also animal species: many Arthropods, but also Vertebrates use dry grasslands and heathlands to feed and reproduce.
And it is fundamental to make people understand that “forest” is not the only habitat type worthy of conservation and protection. Our LIFE will be useful – we hope – also in this sense.
The colours of a dry grassland at the end of May, with the soft indigo of Jasione montana and the bright fuchsia of Silene armeria
Of course, this LIFE is about not only lichens and vascular plants, but it aims at recreating the whole Habitats with their natural whole biodiversity and ecosystem functioning. But I see as a great goal the fact that terricolous lichens are not only considered, but even recognized as a fundamental part of the diversity and functioning of the Habitats the LIFE aims at recreating! My main role in the ideation and writing of the project was exactly to highlight this fundamental role.
As any LIFE project worth of its name, DRYLANDS shall soon have its own website. However, I will update also this blog with the “lichenological” news concerning the advancement of the project, so…stay tuned!